
Scott Solomon
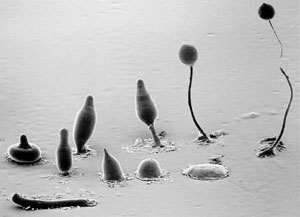
A forest of the fruiting bodies of the social amoeba Dictyselium discoideum. An amoeba that must succeed at both single-celled and multicellular living to pass on its genes, Dicty allows scientists to ask questions about cooperation and cheating in multicellular organisms.
Any multicellular animal, from a blue whale to a human being, poses a special difficulty for the theory of evolution. Most of the cells in its body will die without reproducing, and only a privileged few will pass their genes to the next generation.
How could the extreme degree of cooperation multicellular existence requires ever evolve? Why aren’t all creatures unicellular individualists determined to pass on their own genes?
Joan Strassmann, PhD, and David Queller, PhD, a husband and wife team of evolutionary biologists at Washington University in St. Louis, provide an answer in the Dec. 16 issue of the journal Science. Experiments with amoebae that usually live as individuals but must also join with others to form multicellular bodies to complete their life cycles showed that cooperation depends on kinship.
If amoebae occur in well-mixed cosmopolitan groups, then cheaters will always be able to thrive by freeloading on their cooperative neighbors. But if groups derive from a single cell, cheaters will usually occur in all-cheater groups and will have no cooperators to exploit.
The only exceptions are brand new cheater mutants in all-cooperator groups, and these could pose a problem if the mutation rate is high enough and there are many cells in the group to mutate. In fact, the scientists calculated just how many times amoebae that arose from a single cell can safely divide before cooperation degenerates into a free-for-all.
The answer turns out to be 100 generations or more.
So population bottlenecks that kill off diversity and restart the population from a single cell are powerful stabilizers of cellular cooperation, the scientists conclude.
In other words our liver, blood and bone cells help our eggs and sperm pass on their genes because we passed through a single-cell bottleneck at the moment of conception.
The social amoebae
Wikipedia Commons
Some stages in the life cycles of a social amoeba. When bacteria are scarce, the amoebae send out a distress signal, rush together to form a loose aggregate (second row, right), then a tight aggregate (second row, middle) and then a finger (second row, left). The finger falls over and becomes a slug (front row, far left) that crawls toward heat and light. Once the slug finds a suitable spot, the back end spreads out, raising the front end in the air (the “Mexican hat” at far left). The front end elongates to form a stalk and the back end of the slug flows up the stalk, reorganizing itself at the top into a ball of spores (back row, right).
Queller, the Spencer T. Olin professor, and Strassmann, professor of biology, moved to WUSTL from Rice University this summer, bringing a truckload of frozen spores with them.
Although they worked for many years with wasps and stingless bees, Queller and Strassmann’s current “lab rat” is the social amoeba Dictyostelium discoideum, known as Dicty for short.
The social amoebae can be found almost everywhere; in Antarctica, in deserts, in the canopies of tropical forests, and in Forest Park, the urban park that adjoins Washington University.
The amoebae spend most of their lives as tiny amorphous blobs of streaming protoplasm crawling through the soil looking for E. coli and other bacteria to eat.
Things become interesting when bacteria are scarce and the amoebae begin to starve. They then release chemicals that attract other amoebae, which follow this trail until they bump into one another.
A mound of some 10,000 amoebae forms and then elongates into a slug a few millimeters long that crawls forward (but never backward) toward heat and light.
The slug stops moving when it has reached a suitable place for dispersal, and then the front 20 percent of the amoebae die to produce a sturdy stalk that the remaining cells flow up and there become hardy spores.
Crucially, the 20 percent of the amoebae in the stalk sacrifice their genes so that the other 80 percent can pass theirs on.
When Strassmann and Queller began to work with Dicty in 1998, one of the first things they discovered was that the amoebae sometimes cheat.
Dennis Welker of Utah State University had given them a genetically diverse collection of wild-caught clones (genetically identical amoebae). They mixed amoebae from two clones together and then examined the fruiting bodies to see where the clones ended up. Each fruiting body included cells from both clones, but some clones contributed disproportionately to the spore body. They had cheated.
How can a blob of protoplasm cheat? The answer, it turns out, is many different ways.
“They might,” Queller says, “have a mutation that makes an adhesion molecule less sticky, for example, so that they slide to the back of the slug, the part that forms spores.”
“But there are tradeoffs,” Strassmann says, “because if you’re too slippery, you’ll fall off the slug and lose all the advantages of being part of group.”
Natural born cheaters
Mulling this over, Strassmann and Queller began to wonder if it would be possible to break the social contract among the amoebae by setting up conditions where relatedness was low and each clonal lineage encountered mostly strangers and rarely relatives.

Together with then-graduate student, Jennie Kuzdzal-Fick, they set up an experiment to learn what happened to cheating as heterogeneous (low relatedness) populations of amoebae evolved.
“At the end of the experiment, we assessed the cheating ability of the descendants by mixing equal numbers of descendants and ancestors and checking to see whether the descendants ended up in the stalks or the spores of the fruiting bodies,” Strassmann says.
They found that in nearly all cases, the descendants cheated their ancestors. What’s more, when descendent amoebae were grown as individual clones, about a third of them were unable to form fruiting bodies.
Many of the mutants, in other words, were “obligate” cheaters. Having lost the ability to form their own fruiting bodies, they were able to survive only by freeloading, or taking advantage of the amoebae that had retained the ability to cooperate.
This result, Queller and Strassmann say, shows that cheater mutations that threaten multicellularity occur naturally and are even favored — as long as the population of amoebae remains genetically diverse.
What happens in the wild?
But the scientists were aware that obligate cheaters are either very rare or altogether missing among wild social amoebae. They had not found any obligate cheaters in the more than 2,000 wild clones they have sampled.
They also knew that in the wild, the amoebae in fruiting bodies are close kin, if not clones.
What prevents cooperation in wild populations from degenerating into the laboratory free-for-all? Could the difference be that the amoebae in the laboratory were distant relations and those in the wild are kissing kin?
Suppose, the scientists thought, one amoeba ventured alone into a pristine field of bacteria. As it grew and multiplied, making copies of itself, how long would it take for cheating mutations to appear (what was the mutation rate) and how successfully would these mutations proliferate (how strongly would they be selected)?
To establish the mutation rate, Strassmann and Queller together with graduate student Sara Fox ran what is called a mutation accumulation experiment.
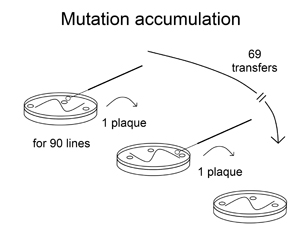
In this experiment, amoebae that mutated didn’t have to compete against amoebae that were faithful replicators. In the absence of selection, all but the most severe mutations were also reproduced and became a permanent part of the lineage’s genome.
The scientists allowed 90 different lines of amoebae to accumulate mutations in this way.
“At the end,” Queller says, “we found that among those 90 lines not a single one had lost the ability to fruit. So that’s almost 100 lines, almost a thousand generations, so 100,000 opportunities to lose fruiting and none of them did.
“That allowed us, using statistics, to put an upper limit on the rate at which mutations turn a cooperator into an obligate cheater,” he says.
The rate was low enough that if fruiting bodies were forming in the wild from amoebae that were all descended from one spore, cheating would never be an issue.
What this has to do with elephants and blue whales
But the scientists were inquisitive enough to ask another, bigger question. They used calculations invented for population genetics to ask how many times the amoeba could divide — theoretically — before cheating became a problem.
What if, they asked, we let an initial single amoebae divide until there were as many of amoebae as there are cells as a fruit fly and then transferred one amoeba and allowed it to divide until the daughter colony reached fruit-fly size, and so on?
What if we let the colonies grow to human size? To elephant size? To blue whale size? Would the cheaters bring down the whale-sized Dicty colony?
The answer, it turned out, was no.
A whale-sized Dicty colony is not the same thing as a whale, but nonetheless the experiments suggest how organisms, over the course of evolution, have sidestepped the cheating trap and maintained the levels of cooperation multicellular bodies demand.
“A multicellular body like the human body is an incredibly cooperative thing,” Queller says, “and sociobiologists have learned that really cooperative things are hard to evolve because of the potential for cheating.
“It’s the single-cell bottleneck that generates high relatedness among the cells that, in turn, allows them to cooperate, ” he says.
Our liver cells have no kick against our sperm or egg cells, in other words, because they’re all nearly genetically identical descendants of a single fertilized egg.